Nature of science:
Developments in scientific research follow improvements in analysis and deduction—improvements in analytical techniques allowing the detection of trace amounts of substances has led to advances in the understanding of plant hormones and their effect on gene expression.
Understandings:
Plant tissues and meristems
Most plants have three basic types of tissues:
Apical meristems
Meristems are often differentiated based on their location within the plant. Apical meristematic tissue, sometimes referred to as primary meristem, occurs at the tips of roots and stems. The apical meristem and the surrounding developing tissue are known as the shoot apex. The shoot apex produces new tissue and causes primary growth through the process of mitosis and cell division Primary growth allows the root to extend throughout the soil. It also allows the stem to grow longer and so increases exposure to light and carbon dioxide. This type of growth results in herbaceous, non-woody stems and roots.
Lateral meristems
Lateral meristems allow the growth in thickness of plants. This is referred to as secondary growth. Most trees and shrubs (woody plants) have active lateral meristems. In fact, these plants have two types of lateral meristem:
Plant Hormones
Plant development involves extensive coordination amongst the individual cells that make up the plant. This requires a means of communication within the plant. Research has shown that there are many factors that affect plant development and growth. They include:
• environmental factors, such as day length and water availability
• receptors, which allow the plant to detect certain environmental factors • the genetic makeup of the plant
• hormones, which are chemical messengers.
Specific cells have proteins (receptors) in their plasma membrane, cytoplasm, or nucleus that allow them to receive different environmental stimuli. Upon reception of a certain stimulus, the protein receptor becomes activated, initiating a metabolic pathway. This pathway often results in the production of a hormone (a chemical messenger). These hormones are produced in very small amounts and have effects in many parts of the plant. The hormones move in the plant through the phloem or from cell to cell. Cells on which a hormone has an effect are referred to as target cells.
Auxins and phototropism
Tropisms are generally defined as growth or movement to directional external stimuli. Tropisms may be positive (towards the stimulus) or negative (away from the stimulus). Common stimuli for plant tropisms include chemicals, gravity, touch, and light. Let’s consider light as a stimulus. Phototropism means plant growth in response to light. Generally, plant stems exhibit positive phototropism, and plant roots demonstrate negative phototropism
Developments in scientific research follow improvements in analysis and deduction—improvements in analytical techniques allowing the detection of trace amounts of substances has led to advances in the understanding of plant hormones and their effect on gene expression.
Understandings:
- Undifferentiated cells in the meristems of plants allow indeterminate growth.
- Mitosis and cell division in the shoot apex provide cells needed for extension of the stem and development of leaves.
- Plant hormones control growth in the shoot apex.
- Plant shoots respond to the environment by tropisms.
- Auxin efflux pumps can set up concentration gradients of auxin in plant tissue.
- Auxin influences cell growth rates by changing the pattern of gene expression.
- Application: Micropropagation of plants using tissue from the shoot apex, nutrient agar gels and growth hormones.
- Application: Use of micropropagation for rapid bulking up of new varieties, production of virus-free strains of existing varieties and propagation of orchids and other rare species.
- Auxin is the only named hormone that is expected.
Plant tissues and meristems
Most plants have three basic types of tissues:
- dermal tissue, which is an outer covering that protects against physical agents and pathogenic organisms, prevents water loss, and may have specialized structures for various purposes
- ground tissue, which consists mostly of thin-walled cells that function in storage, photosynthesis, support, and secretion
- vascular tissue, made up of xylem and phloem that carry out long-distance conduction of water, minerals, and nutrients within the plant, and provide support.
Apical meristems
Meristems are often differentiated based on their location within the plant. Apical meristematic tissue, sometimes referred to as primary meristem, occurs at the tips of roots and stems. The apical meristem and the surrounding developing tissue are known as the shoot apex. The shoot apex produces new tissue and causes primary growth through the process of mitosis and cell division Primary growth allows the root to extend throughout the soil. It also allows the stem to grow longer and so increases exposure to light and carbon dioxide. This type of growth results in herbaceous, non-woody stems and roots.
Lateral meristems
Lateral meristems allow the growth in thickness of plants. This is referred to as secondary growth. Most trees and shrubs (woody plants) have active lateral meristems. In fact, these plants have two types of lateral meristem:
- vascular cambium, which produces secondary vascular tissue and lies between the xylem and the phloem in the vascular bundles, on the inside it produces secondary xylem, which is a major component of wood, and on the outside it produces secondary phloem
- cork cambium, which occurs within the bark of a plant and produces the cork cells of the outer bark.
Plant Hormones
Plant development involves extensive coordination amongst the individual cells that make up the plant. This requires a means of communication within the plant. Research has shown that there are many factors that affect plant development and growth. They include:
• environmental factors, such as day length and water availability
• receptors, which allow the plant to detect certain environmental factors • the genetic makeup of the plant
• hormones, which are chemical messengers.
Specific cells have proteins (receptors) in their plasma membrane, cytoplasm, or nucleus that allow them to receive different environmental stimuli. Upon reception of a certain stimulus, the protein receptor becomes activated, initiating a metabolic pathway. This pathway often results in the production of a hormone (a chemical messenger). These hormones are produced in very small amounts and have effects in many parts of the plant. The hormones move in the plant through the phloem or from cell to cell. Cells on which a hormone has an effect are referred to as target cells.
Auxins and phototropism
Tropisms are generally defined as growth or movement to directional external stimuli. Tropisms may be positive (towards the stimulus) or negative (away from the stimulus). Common stimuli for plant tropisms include chemicals, gravity, touch, and light. Let’s consider light as a stimulus. Phototropism means plant growth in response to light. Generally, plant stems exhibit positive phototropism, and plant roots demonstrate negative phototropism
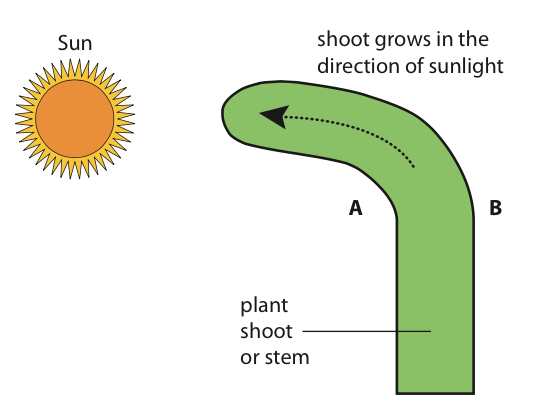
The importance of phototropism to a plant is clear. If an area is crowded with plants, it is essential for seedlings to grow towards the sunlight so that photosynthesis can occur efficiently. Auxins are plant hormones that cause the positive phototropism of plant shoots and seedlings.
Auxins are found in the embryo of seeds, the meristems of apical buds (shoot apex), and young leaves. These hormones only work on plant cells that have auxin receptors. Auxins appear to increase the flexibility of plant cell walls in young developing shoots. This enables cell elongation on the side of the shoot necessary to cause growth towards the light.
This growth response does not appear to be the result of an increased production of auxin on one side of the shoot. Rather, it seems to be caused by a redistribution of available auxin, especially to the side of the stem away from the light source. In the case of phototropism, auxin is actually produced in all the cells in this region of the plant. Auxin efflux pumps (specialized membrane proteins) move the auxins out of the cells closer to the light, using ATP as the energy source. This pumping action creates a high concentration of auxin in the space between the cells. The result is a high concentration of auxin in the intercellular space and a relatively low concentration within the adjacent cell. Because of this, auxin moves down the concentration gradient from the intercellular space into the adjacent cell. The entry of auxin into a cell is called auxin influx. This mechanism of auxin movement between adjacent cells continues until there is a greater concentration of auxin on the stem’s dark side. The result is a greater elongation of cells on the stem side away from the light and, therefore, curvature towards the light source. The specific plant auxin that causes this described action is indoleacetic acid (IAA).
The elongation of the cells is caused by an expansion of the cell walls on the side away from the light source. The key step in this expansion is when auxin combines with a receptor that targets specific transcriptional repressors of auxin-responsive genes.
Auxins are found in the embryo of seeds, the meristems of apical buds (shoot apex), and young leaves. These hormones only work on plant cells that have auxin receptors. Auxins appear to increase the flexibility of plant cell walls in young developing shoots. This enables cell elongation on the side of the shoot necessary to cause growth towards the light.
This growth response does not appear to be the result of an increased production of auxin on one side of the shoot. Rather, it seems to be caused by a redistribution of available auxin, especially to the side of the stem away from the light source. In the case of phototropism, auxin is actually produced in all the cells in this region of the plant. Auxin efflux pumps (specialized membrane proteins) move the auxins out of the cells closer to the light, using ATP as the energy source. This pumping action creates a high concentration of auxin in the space between the cells. The result is a high concentration of auxin in the intercellular space and a relatively low concentration within the adjacent cell. Because of this, auxin moves down the concentration gradient from the intercellular space into the adjacent cell. The entry of auxin into a cell is called auxin influx. This mechanism of auxin movement between adjacent cells continues until there is a greater concentration of auxin on the stem’s dark side. The result is a greater elongation of cells on the stem side away from the light and, therefore, curvature towards the light source. The specific plant auxin that causes this described action is indoleacetic acid (IAA).
The elongation of the cells is caused by an expansion of the cell walls on the side away from the light source. The key step in this expansion is when auxin combines with a receptor that targets specific transcriptional repressors of auxin-responsive genes.
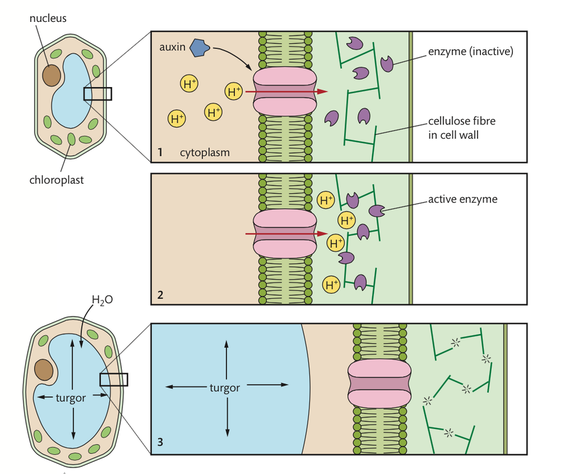
The sequence of events in the stem causing it to bend towards a light source is as follows.
- Auxin is produced by all cells in the stem on the side towards the light source.
- Auxin moves by efflux pump action into the nuclei of cells on the side of the stem
opposite the light.
- The auxin and a receptor in the nuclei form a complex that activates a proton
(hydrogen ion) pump. Present research indicates that auxin itself also has an effect on
the protein pump of the stem cell membranes as indicated in Figure 9.19.
- The proton pump moves hydrogen ions into the spaces of the cell wall.
- These hydrogen ions cause a drop in pH, resulting in the hydrogen bonds between
cellulose fibres of the cell wall breaking.
- This results in the elongation of the cells on the side away from the light.
Auxins are a very complex set of hormones that have many functions within a plant. As well as the effect they have in phototropism, auxins are also involved in:
• stimulation of cell division in most meristematic tissue
• differentiation of xylem and phloem
• development of lateral roots in tissue cultures
• suppression of lateral bud growth when present in the apical bud • stimulation of growth of flower parts
• induction of fruit production without pollination.
Theory of Knowledge
Plants communicate chemically both internally and externally. To what extent can plants be said to have language?
In order to answer this question, first, we need to know what the meaning of language is:
"The system of communication used by a particular community or country"
The "community" mentioned above could be used to make reference to all the structures within the plant's organism, such as living and functioning cells. For language to exist, there needs to be some sort of communication, and as the question states it, plants communicate chemically internally and externally.
In brief, plants do indeed have their own "language"; communicating chemically is their way of painting their wellbeing within themselves and the environment that surrounds them.
Plants communicate chemically both internally and externally. To what extent can plants be said to have language?
In order to answer this question, first, we need to know what the meaning of language is:
"The system of communication used by a particular community or country"
The "community" mentioned above could be used to make reference to all the structures within the plant's organism, such as living and functioning cells. For language to exist, there needs to be some sort of communication, and as the question states it, plants communicate chemically internally and externally.
In brief, plants do indeed have their own "language"; communicating chemically is their way of painting their wellbeing within themselves and the environment that surrounds them.
